Visión
general de la fotosíntesis: sus etapas
Los
organismos fotosintéticos productores de O2 usan energía lumínica,
CO2 y agua para producir la materia orgánica necesaria para su
alimentación. El O2 que liberan se forma con átomos provenientes
del agua.
La
fotosíntesis se realiza en dos etapas: la lumínica, en la que se
utiliza la energía de la luz para sintetizar ATP y NADPH, y la
fijadora de carbono, que utiliza los productos de la primera etapa
para la producción de azúcares.
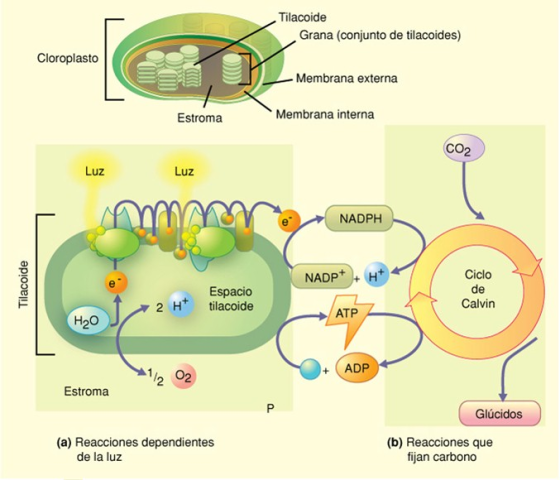
La
fotosíntesis ocurre en dos etapas: las reacciones dependientes de la
luz y las reacciones que fijan carbono. (a) En las reacciones
dependientes de la luz, la absorción de la energía lumínica por
las moléculas de clorofila a en la membrana del tilacoide inicia un
transporte de electrones y la formación de un gradiente de protones
a partir del cual se produce ATP. Durante este proceso, la molécula
de agua se escinde y se liberan moléculas de oxígeno gaseoso. Los
electrones son finalmente absorbidos por el NADP+ y se forma NADPH.
(b) En las reacciones que fijan carbono, que ocurren en la estroma
del cloroplasto, se sintetizan glúcidos a partir del CO2 y el
hidrógeno que transporta el NADPH. Este proceso utiliza la energía
del ATP y el NADPH producidos en la etapa dependiente de la luz y,
como veremos más adelante, implica una serie de reacciones que
constituyen el ciclo de Calvin.
La
fotosíntesis se realiza en los cloroplastos: los tilacoides
En
los eucariontes, la fotosíntesis se realiza en los cloroplastos,
organelas que poseen una membrana externa y otra interna. La membrana
interna rodea una solución densa, la estroma, donde se encuentran
las membranas tilacoides, que tienen forma de sacos aplanados
dispuestos en forma apilada. Las reacciones de la etapa lumínica
ocurren en los sacos tilacoides y las que fijan el carbono, en la
estroma.
Los
sacos tilacoides de los procariontes fotosintéticos pueden formar
parte de la membrana celular, estar aislados en el citoplasma o
constituir una estructura compleja de la membrana interna.
La
naturaleza de la luz
El
modelo ondulatorio de la luz permite a los físicos describir
matemáticamente ciertos aspectos de la luz y el modelo fotónico
permite otro tipo de cálculos y predicciones matemáticas. Estos dos
modelos ya no se consideran opuestos uno al otro, sino
complementarios, en el sentido de que es necesaria una síntesis de
ambos para una descripción completa del fenómeno que conocemos como
luz.
Los
sistemas vivos absorben la energía lumínica mediante el uso de
pigmentos. Los organismos fotosintéticos tienen distintos tipos de
pigmentos: la clorofila, que se encuentra en los sacos tilacoides,
los carotenoides y las ficobilinas. Existen diferentes tipos de
clorofila: la clorofila a, que colecta energía luminosa y está
involucrada en la transformación de energía lumínica en química;
la clorofila b, presente en las plantas y las algas verdes, y la
clorofila c de las algas marrones.
La
correspondencia entre el espectro de absorción de las clorofilas a y
b y el espectro de absorción de la fotosíntesis indica una estrecha
relación entre ésta y aquéllas (en ambos casos se observan dos
picos, uno en la zona del rojo y otro en la del azul). Los
carotenoides absorben en forma muy eficiente longitudes de onda que
no son absorbidas por la clorofila.
El
transporte de electrones: los fotosistemas y la ATP sintetasa
Los
organismos fotosintéticos poseen dos fotosistemas, cada uno formado
por una antena colectora de luz y un centro de reacción fotoquímico
que incluye una molécula de clorofila a. Ambos fotosistemas se
diferencian por el pico de absorción de la clorofila: el Fotosistema
I lo presenta a 700 nm; el Fotosistema II, a 680 nm.
En
un flujo no cíclico de electrones, los dos fotosistemas trabajan en
forma simultánea y continua. Así se produce un flujo permanente de
electrones desde el agua al Fotosistema II, de éste al Fotosistema I
y de este último al NADP+.
Durante
el transporte de electrones, los protones presentes en la estroma son
enviados al espacio intertilacoide, creando un gradiente cuya energía
se utiliza para sintetizar ATP. La síntesis de ATP a partir de
energía lumínica se conoce como fotofosforilación.
Cuando
los dos fotosistemas trabajan en forma independiente, se forma un
flujo cíclico de electrones. En este caso no se forma NADPH, pero se
sintetiza ATP. Es una ruta alternativa que permite regular la
cantidad de NADPH y ATP formados en presencia de luz y,
probablemente, aumenta la eficiencia en la formación de ATP cuando
coexiste con el flujo no cíclico de electrones.
Los fotosistemas:

La
energía lumínica atrapada en la molécula reactiva de la clorofila
a del Fotosistema II lanza los electrones a un nivel de energía
superior. Estos electrones son reemplazados en la molécula de
clorofila a por electrones que provienen indirectamente de moléculas
de agua que se escinden liberando además protones (H+) y gas
oxígeno. Los electrones pasan desde el aceptor de electrones
primario, a lo largo de una cadena de transporte de electrones, a un
nivel de energía inferior, el centro de reacción del Fotosistema I.
A medida que pasan a lo largo de esta cadena de transporte de
electrones, se forma un gradiente de protones a partir del cual se
sintetiza ATP. La energía lumínica absorbida por el Fotosistema I
lanza los electrones a otro aceptor primario. Desde este aceptor, los
electrones son transferidos mediante otros transportadores al NADP+ y
se forma NADPH. Los electrones eliminados del Fotosistema I son
reemplazados por los del Fotosistema II. El ATP y el NADPH
representan la ganancia neta de las reacciones que capturan energía.
La
fotofosforilación:
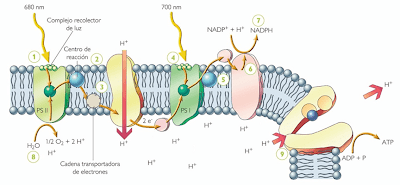
Moléculas
y complejos moleculares que participan de las reacciones directamente
dependientes de la luz. Entre ellos, se distinguen los pigmentos, los
transportadores de electrones, los Fotosistemas I y II y ciertas
enzimas como las ATP sintetasas. La disposición particular de estas
moléculas en la membrana tilacoide hace posible la síntesis
quimiosmótica del ATP durante la fotofosforilación. En este
proceso, los electrones de la molécula reactiva de clorofila a del
Fotosistema II son impulsados a niveles energéticos superiores por
la energía lumínica. A medida que descienden por una cadena de
transportadores de electrones hacia la molécula reactiva de
clorofila a del Fotosistema I, la energía que liberan se usa para
bombear protones (H+) desde la estroma al espacio tilacoide. Esto
crea un gradiente de protones. Cuando los protones se mueven a favor
del gradiente a través del complejo de la ATP sintetasa, desde el
espacio tilacoide a la estroma del cloroplasto, el ADP se fosforila a
ATP.
Las
reacciones que fijan carbono
El
ATP y el NADPH formados durante el transporte de electrones se
utilizan en la reducción del CO2 a glucosa. La incorporación de CO2
en compuestos orgánicos se conoce como fijación del carbono y
ocurre en forma cíclica (ciclo de Calvin). En las plantas verdes, el
CO2 llega a las células fotosintéticas a través de aberturas
especializadas llamadas estomas.
El
ciclo de Calvin comienza con la unión del CO2 a una molécula de
cinco carbonos (ribulosa bifosfato) que luego se divide en dos
moléculas de tres carbonos (fosfoglicerato). Cada seis vueltas del
ciclo se introducen seis moléculas de CO2 y se producen dos
moléculas de un azúcar de tres carbonos (gliceraldehído fosfato).
Las
plantas poseen un mecanismo de control que evita que el ciclo de
Calvin ocurra durante la noche. La luz lo estimula indirectamente y
las reacciones de fijación de carbono son inhibidas en la oscuridad.
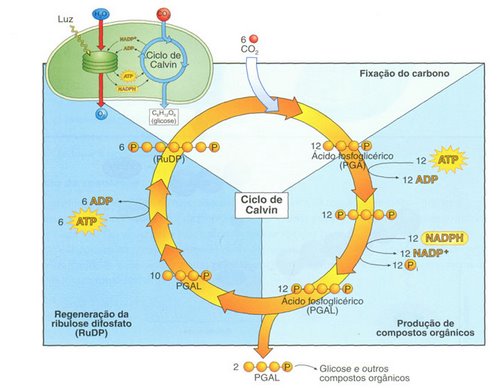
En
cada "vuelta" completa del ciclo ingresa una molécula de
CO2. Aquí se resumen seis ciclos, el número requerido para elaborar
dos moléculas de gliceraldehído fosfato, que equivalen a un azúcar
de seis carbonos. Se combinan seis moléculas de ribulosa bifosfato
(RuBP), un compuesto de cinco carbonos, con seis moléculas de CO2 y
se producen seis moléculas de un intermediario inestable que pronto
se escinde en doce moléculas de fosfoglicerato, un compuesto de tres
carbonos. Estos últimos se reducen a doce moléculas de
gliceraldehído fosfato. Diez de estas moléculas de tres carbonos se
combinan y se regeneran para formar seis moléculas de cinco carbonos
de RuBP. Las dos moléculas "extra" de gliceraldehído
fosfato representan la ganancia neta del ciclo de Calvin. Estas
moléculas son el punto de partida de numerosas reacciones que pueden
implicar, por ejemplo, la síntesis de glúcidos, aminoácidos y
ácidos grasos. La energía que impulsa al ciclo de Calvin proviene
del ATP y el NADPH producidos por las reacciones de captura de
energía en la primera etapa de la fotosíntesis.
La
fotorrespiración ocurre cuando la concentración de CO2 en la hoja
es baja en relación con la de O2. Consiste en la oxidación de la
ribulosa bifosfato, con formación de CO2 y agua. Es un proceso que
disminuye la eficiencia fotosintética de las plantas.
En
las células del mesófilo de las plantas C4, el CO2 se une a un
compuesto de tres carbonos (fosfoenolpiruvato), formando oxalacetato.
Este último se convierte en malato y pasa a zonas más profundas de
la hoja, donde libera CO2 que ingresa en el ciclo de Calvin. Este
proceso, que involucra gasto de energía, representa una adaptación
a las sequías y a intensidades lumínicas y temperaturas altas.

El
CO2 se fija primero en las células del mesófilo como ácido
oxalacético. La PEP carboxilasa, a diferencia de la RuBP
carboxilasa, es incapaz de incorporar O2. Aun con concentraciones muy
bajas de CO2 y en presencia de abundante O2, la enzima trabaja
rápidamente uniendo el CO2 al PEP. Comparada con la RuBP
carboxilasa, en presencia de O2 la PEP carboxilasa fija el CO2 más
rápido y en concentraciones menores, manteniendo baja la
concentración de CO2 dentro de las células cercanas a la superficie
de la hoja. El ácido oxalacético se transforma en ácido málico
que es transportado a las células de la vaina, donde libera CO2. El
CO2 así formado entra en el ciclo de Calvin. El ácido málico se
transforma en pirúvico que regresa a la célula del mesófilo, donde
es fosforilado a PEP.
En
plantas de ambientes secos existe una vía metabólica llamada
fotosíntesis CAM. La fijación de CO2 ocurre durante la noche y con
él se forma malato, que se almacena en las vacuolas. Durante el día,
el malato es liberado, se descarboxila y el CO2 ingresa en el ciclo
de Calvin.
Utilización
de los productos de la fotosíntesis
El
gliceraldehído fosfato producido por el ciclo de Calvin se integra
en glucosa o fructosa. Las células vegetales usan estas sustancias
para elaborar almidón, celulosa y sacarosa; las células animales
las usan para elaborar glucógeno. Todas las células utilizan
azúcares para la elaboración de otros carbohidratos, lípidos y
aminoácidos. Además, la oxidación del carbono fijado es la fuente
de energía del ATP en todas las células heterótrofas.
El
balance entre la fotosíntesis y la respiración
En
las plantas, la fotosíntesis y la respiración ocurren en forma
simultánea. La intensidad lumínica a la cual se igualan sus
velocidades es el punto de compensación para la luz. La
concentración de CO2 a la cual se igualan es el punto de
compensación para el CO2. Por debajo de estos puntos de
compensación, la respiración excede a la fotosíntesis y la planta
no crece. Como muchos órganos vegetales no fotosintetizan, para que
una planta se mantenga y crezca, la fotosíntesis debe exceder
largamente la tasa de respiración.
No hay comentarios. :
Publicar un comentario