lunes, 19 de diciembre de 2016
El flujo de información genética: los caminos del DNA a la proteína
A
partir de la década de 1940, el concepto de gen evolucionó desde la
formulación "un gen: una enzima" a "un gen: una
cadena polipeptídica". Sin embargo, ninguna de estas
definiciones se ajusta por completo al concepto actual de gen.
El
flujo de información dentro de la célula
El
"dogma central de la biología", definido en 1957 por
Francis Crick, establece que la información genética fluye en el
siguiente sentido: DNA → RNA→ proteínas. Esto es verdad en la
mayoría de los casos; sin embargo, el material genético de algunos
virus está formado por RNA que luego es usado como molde para
producir DNA.
El
código genético
El
código genético consiste en la asignación de tripletes de
nucleótidos (codones) en el RNA mensajero (mRNA) a cada uno de los
aminoácidos que formarán una cadena polipeptídica.
Existen 64 combinaciones posibles de codones. El código es redundante, porque los 20 aminoácidos usualmente presentes en los seres vivos son codificados por 61 de estas combinaciones. Los tres codones restantes actúan como señales de terminación de la traducción.
Con muy pocas excepciones, el código genético es el mismo en casi todos los seres vivos.
Existen 64 combinaciones posibles de codones. El código es redundante, porque los 20 aminoácidos usualmente presentes en los seres vivos son codificados por 61 de estas combinaciones. Los tres codones restantes actúan como señales de terminación de la traducción.
Con muy pocas excepciones, el código genético es el mismo en casi todos los seres vivos.
La
transcripción: del DNA al RNA
La
transcripción es el proceso de síntesis de RNA a partir de DNA.
Sigue el mismo principio de apareamiento de bases que la replicación
del DNA, pero se reemplaza la timina por el uracilo. En cada
transcripción, sólo una de las cadenas del DNA se transcribe. La
RNA polimerasa cataliza la adición de ribonucleótidos al extremo 3´
de la cadena de RNA, de modo que esta última es antiparalela a la
cadena molde de DNA.

En
la región del promotor, punto de unión de la enzima RNA polimerasa,
la doble hélice de DNA se abre y, a medida que la RNA polimerasa
avanza a lo largo de la molécula de DNA, se separan las dos cadenas.
Los ribonucleótidos, que constituyen los bloques estructurales, se
ensamblan en la dirección 5' a 3' a medida que la enzima lee la
cadena molde de DNA. Nótese que la cadena de RNA recién sintetizada
es complementaria, no idéntica, a la cadena molde a partir de la
cual se transcribe; su secuencia, sin embargo, es idéntica a la
cadena codificante de DNA (no transcrita), excepto por un detalle: en
el RNA, la timina (T) se reemplaza por uracilo (U). El RNA recién
sintetizado se separa de la cadena molde de DNA.
La
RNA polimerasa no necesita un cebador para iniciar la síntesis. Se
une al DNA en una secuencia específica, el promotor, que define el
punto de inicio de la transcripción y su dirección.
En los procariontes, el proceso de transcripción continúa hasta que la polimerasa encuentra una secuencia que constituye la señal de terminación. En los eucariontes, el proceso termina cuando el RNA es cortado en una secuencia específica. Al finalizar la transcripción, la RNA polimerasa se detiene y libera la cadena molde de DNA y el mRNA sintetizado.
En los eucariontes, los transcritos primarios sufren diversas modificaciones durante la transcripción. Entre ellas se encuentran la adición del CAP, la poliadenilación y el splicing. Este último proceso consiste en el corte y la eliminación de ciertas secuencias, los intrones, y el posterior empalme de las secuencias restantes, los exones. Sólo los exones forman parte del mRNA maduro. Un mismo transcrito primario puede ser procesado por splicing de distintas maneras. Este empalme alternativo permite que una molécula de mRNA inmadura pueda originar diferentes moléculas de mRNA maduro.
En los procariontes, el proceso de transcripción continúa hasta que la polimerasa encuentra una secuencia que constituye la señal de terminación. En los eucariontes, el proceso termina cuando el RNA es cortado en una secuencia específica. Al finalizar la transcripción, la RNA polimerasa se detiene y libera la cadena molde de DNA y el mRNA sintetizado.
En los eucariontes, los transcritos primarios sufren diversas modificaciones durante la transcripción. Entre ellas se encuentran la adición del CAP, la poliadenilación y el splicing. Este último proceso consiste en el corte y la eliminación de ciertas secuencias, los intrones, y el posterior empalme de las secuencias restantes, los exones. Sólo los exones forman parte del mRNA maduro. Un mismo transcrito primario puede ser procesado por splicing de distintas maneras. Este empalme alternativo permite que una molécula de mRNA inmadura pueda originar diferentes moléculas de mRNA maduro.

La
información genética codificada en el DNA se transcribe a una copia
de RNA (transcripto primario). Esta copia se modifica en forma
cotranscripcional con la adición del casquete 5' (CAP), el corte de
los intrones y el empalme de los exones (splicing) y, finalmente con
la adición de la cola de poli-A. A ambos extremos del mensajero hay
secuencias no traducibles, denominadas extremos 5´UTR (región no
traducible que abarca desde el CAP hasta el codón de iniciación) y
extremos 3´UTR (región no traducible que abarca desde el codón de
terminación hasta la cola de PoliA). En esta figura, el splicing se
produce luego de la adición de la cola de poli-A, sin embargo,
muchas veces el proceso de corte y empalme ocurre antes de que haya
concluido la transcripción. El mRNA maduro luego se dirige
al citoplasma,
donde se traduce a proteínas.
En
el ciliado de agua dulce Tetrahymena, el intrón inmaduro actúa como
catalizador de la escisión, produciendo un empalme autocatalítico.
A este RNA con función de enzima se lo llama ribozima.
La
traducción: del RNA al polipéptido
La
traducción es la conversión de la secuencia de nucleótidos del RNA
en la secuencia de aminoácidos de un polipéptido. En este proceso
participan los mRNA, los RNA ribosómicos (rRNA) y los RNA de
transferencia (tRNA).
Los
ribosomas están formados por rRNA y proteínas. Cada uno está
formado por dos subunidades de diferente tamaño que, además, en los
procariontes son más pequeñas que en los eucariontes.
El
mRNA y el tRNA iniciador se unen a la subunidad ribosómica menor.
Luego se les une la subunidad mayor y cataliza la unión peptídica
entre aminoácidos. En la subunidad mayor existen tres sitios a los
que se une el tRNA: el sitio A (aminoacílico), el sitio P
(peptidílico) y el sitio E (de salida).
Los
tRNA son moléculas pequeñas, con una estructura secundaria
semejante a la hoja de un trébol, que presentan dos sitios de unión.
Uno de ellos es el anticodón, que se aparea con el codón del mRNA.
El otro sitio, ubicado en el extremo 3´, se acopla a un aminoácido
particular en forma muy específica. Así, los tRNA permiten la
alineación de los aminoácidos de acuerdo con la secuencia de
nucleótidos del mRNA.
El
grupo de enzimas aminoacil-tRNA sintetasas catalizan la unión entre
el aminoácido y el tRNA y forman el complejo aminoacil-tRNA. Este
complejo se une a la molécula de mRNA, apareando el anticodón con
el codón del mRNA en forma antiparalela. Así, el tRNA coloca al
aminoácido específico en su lugar. El enlace entre el aminoácido y
el tRNA se rompe cuando se forma el enlace entre el aminoácido
recién llegado y el último de la cadena polipeptídica en
crecimiento.
En
los procariontes, el proceso de traducción comienza antes de que
haya finalizado el de transcripción. En los eucariontes, ambos
procesos están separados en el tiempo y en el espacio: la
transcripción ocurre en el núcleo y la traducción, en el
citoplasma.
Tanto
en procariontes como en eucariontes, la síntesis de polipéptidos
ocurre en tres etapas: iniciación, elongación y terminación.
Hacia
el final del mRNA hay un codón que actúa como señal de
terminación. No existe ningún tRNA que tenga un anticodón que se
aparee con este codón. Existen, en cambio, factores de liberación
que se unen al codón de terminación y provocan la separación del
polipéptido y el tRNA. Finalmente, las dos subunidades ribosómicas
también se separan.
Las
proteínas "chaperonas" ayudan a las cadenas polipeptídicas
a plegarse. Finalizado este proceso, las nuevas proteínas viajan al
medio extracelular o a los distintos compartimientos celulares, según
el tipo de señales que posean.
Una
redefinición de las mutaciones
Una
mutación es un cambio en la secuencia o en el número de nucleótidos
en el DNA de una célula. Sólo las mutaciones que ocurren en los
gametos se transmiten a la descendencia. Las mutaciones puntuales
implican la sustitución de un nucleótido por otro. La adición o la
sustracción (deleción) de nucleótidos provoca el corrimiento del
marco de lectura y, por consiguiente, la aparición de una proteína
nueva que casi siempre resulta defectuosa.
Una
revisión del concepto de gen
Actualmente
se considera que un gen es un segmento de DNA que se encuentra a
continuación de un promotor y que puede ser transcrito por una RNA
polimerasa, originando un RNA funcional.
Los experimentos de Mendel y el nacimiento de la genética
Las
contribuciones de Mendel
A mediados del siglo XIX, Gregor Mendel realizó cruzamientos
experimentales con variedades de Pisum sativum (arveja común). Usó
formas puras que poseían características claramente diferentes y no
cambiaban de una generación a otra.
Al cruzar dos variantes de una misma característica, Mendel observó
que en la primera generación (F1) todos los individuos presentaban
sólo una de las variantes, a la que Mendel llamó variante
dominante. En la siguiente generación (F2), obtenida por
autopolinización de la F1, reaparecía la variante ausente en la
primera generación y Mendel la llamó variante recesiva. En la F2,
la proporción entre variante dominante y variante recesiva era
aproximadamente 3:1.
Según la "primera ley de Mendel", o principio de
segregación, cada individuo lleva un par de factores hereditarios
para cada característica. Los miembros del par segregan durante la
formación de los gametos. Los factores hereditarios de Mendel
coinciden con el actual concepto de gen.
El principio de segregación

A
partir de un cruzamiento entre plantas de la generación P, una
planta de guisante homocigótica para el alelo dominante (BB) y la
otra homocigótica para el alelo recesivo (bb), se obtienen las
generaciones F1 y F2. El fenotipo de la progenie -la generación F1-
es púrpura, pero su genotipo es Bb. La F1 heterocigótica produce
cuatro tipos de gametos: masculinos B, femeninos B, masculinos b y
femeninos b, en proporciones iguales. Cuando esta planta se
autopoliniza, los gametos masculinos y los femeninos, B y b, se
combinan al azar y forman, en promedio 1/4 BB (púrpura), 2/4 (o 1/2)
Bb (púrpura) y 1/4 bb (blanco), lo que significa una relación
genotípica de 1:2:1. Esta relación genotípica da cuenta de la
relación fenotípica: tres dominantes (púrpura) a un recesivo
(blanco), que se expresa como 3:1.
Los alelos son variantes de un mismo gen presentes en una población.
Los dominantes se representan con letras mayúsculas y los recesivos
con minúsculas.
Los individuos diploides que tienen dos alelos iguales de un gen
determinado son homocigóticos para ese gen; los individuos cuyos
alelos son distintos son heterocigóticos para el gen en cuestión.
Los alelos dominantes se expresan en la condición homocigota y en la
heterocigota, mientras que los alelos recesivos lo hacen sólo en la
homocigota.
El genotipo es la constitución genética de un individuo. El
fenotipo, resultado de la interacción entre el genotipo y el
ambiente, es su apariencia externa.
Al analizar simultáneamente la herencia de dos características,
Mendel observó que la F1 era homogénea, y que en la F2 aparecían
los cuatro fenotipos posibles en una proporción 9:3:3:1 (doble
dominante:dominante/recesivo:recesivo/dominante:doble recesivo).
La "segunda ley de Mendel", o principio de distribución
independiente, establece que durante la formación de los gametos,
cada par de alelos segrega independientemente de los otros pares.
El
principio de la distribución independiente

Una
planta homocigótica para semillas redondas (RR) y amarillas (AA) se
cruza con una planta de semillas rugosas (rr) y verdes (aa). Toda la
generación Fl tiene semillas redondas y amarillas (RrAa). En la F2,
de las 16 combinaciones posibles en la progenie, 9 muestran las dos
variantes dominantes (RA, redonda y amarilla), 3 muestran una
combinación de dominante y recesivo (Ra, redonda y verde), 3
muestran la otra combinación (rA, rugosa y amarilla) y 1 muestra las
dos recesivas (ra, rugosa y verde). Esta distribución 9:3:3:1 de
fenotipos es el resultado esperado de un cruzamiento en el que
intervienen dos características que se distribuyen
independientemente, cada una con un alelo dominante y uno recesivo en
cada uno de los progenitores.
Sobre
genes y cromosomas
Sutton supuso que los elemente descritos por Mendel -que hoy
conocemos como genes- están en los cromosomas y que los alelos -las
formas alternativas de un gen- se encuentran en cromosomas homólogos.
Cuando se separan los cromosomas homólogos durante la meiosis I,
también se separan los alelos de cada gen y cuando los gametos se
fusionan durante la fecundación, se forman nuevas combinaciones de
alelos.
A comienzos del siglo XX, Hugo de Vries descubrió que las variantes
alélicas se originaban a partir de cambios repentinos en el material
hereditario. A esos cambios los llamó mutaciones.
Los genes que se encuentran en el cromosoma X corresponden a
características ligadas al sexo. En las especies animales cuyos
machos tienen un sistema de cromosomas sexuales XY, las proporciones
fenotípicas de las características ligadas al sexo en la F2 no
cumplen la primera "ley de Mendel".
La segunda "ley de Mendel" tiene validez para genes que se
encuentran en distintos cromosomas o lo suficientemente alejados unos
de otros. El entrecruzamiento entre cromosomas homólogos permite que
los genes se separen y se recombinen. Los genes ligados son aquellos
que, por encontrarse próximos, tienden a segregar juntos.
El lugar que ocupan los genes en los cromosomas se denomina locus.
Los alelos de un mismo gen ocupan el mismo locus en los cromosomas
homólogos.
Los genes están dispuestos linealmente en los cromosomas. Cuanto
menor es la distancia entre ellos, menor es el porcentaje de
recombinación. La frecuencia de recombinación permite conocer el
orden y la distancia de los genes en los cromosomas.
Entrecruzamiento

La
posibilidad de recombinación en cualquier punto entre B y D es mayor
que entre B y C, simplemente porque la distancia entre B y D es mayor
y, en consecuencia, hay más lugar (y por tanto mayor probabilidad)
para que ocurra el entrecruzamiento.
Ampliando
el concepto de gen
En el fenómeno de dominancia incompleta, el cruzamiento de dos
organismos homocigotos (uno dominante y el otro recesivo) origina una
F1 homogénea, pero con un fenotipo intermedio al de ambos
progenitores. En el caso de la codominancia, los individuos
heterocigotos de la F1 expresan en forma simultánea los dos
fenotipos de los progenitores.
Dominancia
incompleta

Un
cruzamiento entre una planta Antirrhinum con flores rojas (R1R1) y
una con flores blancas (R2R2). Este cruzamiento se parece mucho al
realizado entre una planta de guisante de flores púrpura y otra de
flores blancas que se muestra en la figura 8-6, pero hay una
diferencia significativa. En este caso, ningún alelo es dominante.
La flor del heterocigoto presenta un fenotipo intermedio entre los
dos colores.
Se denomina alelos múltiples a la existencia en la población de más
de dos variantes alélicas de un mismo gen. Tales variantes presentan
entre sí diferentes relaciones de dominancia.
Además de las interacciones entre los alelos de un mismo gen,
existen interacciones entre los alelos de genes diferentes. Como
resultado de estas interacciones, en las características
determinadas por más de un gen aparecen diferentes fenotipos.
Epistasis
en arvejillas de olor

Bateson
y sus colegas encontraron un ejemplo claro de interacción génica
epistática en la arvejilla de olor Lathyrus odoratus. Cuando
cruzaron dos variedades puras obtenidas en forma independiente, ambas
de flores blancas, encontraron que toda la progenie de F1 se
autopolinizaran, de 651 plantas que florecieron en la generación F2,
382 tenían flores púrpura y 269, flores blancas. Si examinamos
cuidadosamente estas proporciones, se verá que se ajustan a una
relación 9:7. ¿Cómo podemos explicar este resultado? Recordemos
que en un cruzamiento en el que se analizan dos genes cuyos alelos se
distribuyen independientemente se espera que la relación fenotípica
sea 9:3:3:1. La explicación en este caso es que dos genes diferentes
tienen efecto sobre el color de las flores. Se representan los alelos
de estos genes como C, c, P y p. La proporción fenotípica 9/16 de
F2 muestra los efectos de los dos alelos dominantes (C y P). La
proporción 7/16 reúne al resto de la progenie. Sólo una planta que
haya recibido los alelos dominantes de ambos genes (o sea, el alelo P
y el alelo C) es capaz de producir pigmento púrpura. En este caso,
cualquier gen en la condición homocigoto recesivo es epistático u
oculta el efecto del otro gen. Cuando el gen C es homocigoto recesivo
(cc), las flores son blancas aunque esté presente un alelo P
dominante (como en los fenotipos ccPp y ccPP). De modo semejante,
cuando el gen P es homocigoto recesivo (pp), las flores también son
blancas (como en los fenotipos Ccpp y CCpp).
La epistasis consiste en el enmascaramiento del efecto de un gen por
parte de otro gen. Cuando ocurre esto, las proporciones mendelianas
no se cumplen.
La pleiotropía es el caso en que un gen afecta a más de una
característica.
La herencia poligénica es aquella en la que el fenotipo es el
resultado de la acción acumulativa de los efectos combinados de
muchos genes. Estas características presentan una variación
continua.
La expresividad se refiere a la variación o proporcionalidad en la
expresión de un genotipo; la penetrancia, a fenotipos cuya
frecuencia no coincide con la esperada. Los términos expresividad y
penetrancia se utilizan para referirse a casos en los que el fenotipo
asociado a un gen depende de factores ambientales o desconocidos.
Alteraciones
cromosómicas
Los cromosomas pueden sufrir cambios que afectan su número
(alteraciones numéricas) o su estructura (alteraciones
estructurales). Estos cambios pueden alterar la viabilidad o la
fertilidad de un organismo, o mantenerse como parte de la
variabilidad genética de la población. Se consideran mutaciones y
ocurren durante la mitosis o la meiosis.
La reproducción celular
La
distribución de la información genética
La
división celular permite la reproducción de los organismos
unicelulares y pluricelulares. En estos últimos posibilita, además,
el desarrollo de un individuo a partir de una única célula y la
reparación de los tejidos dañados.
En
los procariontes y los eucariontes, los cromosomas se duplican antes
de la división celular. Luego se distribuyen entre las células
hijas de tal manera que se produce una distribución equitativa del
material hereditario. En los eucariontes existen dos tipos de
división celular: la mitosis y la meiosis.
La
vida de una célula: el ciclo celular
El
ciclo celular es la sucesión de fases de crecimiento y división que
ocurren en la vida de una célula. En él se pueden reconocer tres
fases: interfase, mitosis y citocinesis.
El
ciclo celular:

La
división celular, constituida por la mitosis (cariocinesis o
división del núcleo) y la citocinesis (división del citoplasma),
ocurre después de completarse las tres fases preparatorias de la
interfase: fases G1, S y G2.
La
interfase abarca tres etapas: G1, S y G2. Durante G1, la célula
crece y se duplican las organelas; en las células animales, los
centríolos empiezan a duplicarse. En la etapa S se duplican el DNA y
sus proteínas asociadas. En G2 comienzan a ensamblarse las
estructuras relacionadas con la división celular, los cromosomas se
condensan y los centríolos terminan de duplicarse.
El
ciclo celular está regulado por estímulos externos e internos. La
falta de nutrientes, los cambios de temperatura y de pH, y la
presencia de células contiguas pueden detener la división celular,
mientras que ciertas hormonas y factores de crecimiento la estimulan.
La regulación interna es realizada mediante la fosforilación y la
degradación de complejos proteicos llamados Cdk-ciclinas, formados
por una subunidad reguladora (la ciclina) y otra catalítica (la
cinasa). La actividad de estos complejos determina si el ciclo
celular avanza o se detiene.
La
división del núcleo y del citoplasma: mitosis y citocinesis
La
mitosis es un proceso continuo, en el que se reconocen cuatro fases:
profase, metafase, anafase y telofase.
Al
comienzo de la mitosis, cada cromosoma está formado por dos
cromátidas hermanas y se encuentra totalmente condensado. El huso
mitótico está armado y las fibras cinetocóricas están unidas a
los centrómeros de los cromosomas. Las fibras del huso separan las
cromátidas hermanas, que son conducidas a polos opuestos de la
célula. Así se asegura la distribución equitativa de la
información genética entre las dos células hijas.
La
citocinesis divide a la célula madre en dos hijas casi iguales. Cada
una de ellas recibe un juego completo de cromosomas y alrededor de la
mitad del citoplasma, las organelas y las macromoléculas de la
célula madre.
Senescencia:
el envejecimiento de una célula
El
número de divisiones de las células eucariontes en cultivo
disminuye con el tiempo y está correlacionado con el acortamiento
progresivo de los telómeros. Finalmente, las células entran en un
estado de senescencia, que se caracteriza por la ausencia de división
celular.
El
proceso de muerte celular: apoptosis versus necrosis
La
apoptosis es un proceso de muerte celular programada genéticamente.
En los vertebrados, controla el número de neuronas durante el
desarrollo del sistema nervioso, elimina células defectuosas y da
forma a los órganos en desarrollo. Junto con la mitosis, modela las
formas de los organismos.
Las
caspasas son enzimas que degradan las proteínas de la lámina
nuclear y del citoesqueleto, y provocan la apoptosis. Su actividad
está controlada por otras proteínas que, a su vez, responden a
factores extracelulares.
La
necrosis es un tipo de muerte celular no controlada. Suele producir
la hinchazón y el estallido de las células.
La
división celular: un modo de reproducción de un organismo
En
los organismos unicelulares, la división celular está asociada con
la reproducción y permite la aparición de dos réplicas exactas de
cada individuo.
Hacia
la reproducción sexual
La
reproducción sexual ocurre en la mayoría de los eucariontes.
Requiere dos progenitores y siempre involucra dos procesos: la
meiosis y la fecundación.
Células
haploides, diploides y poliploides: distinto número de dotaciones
cromosómicas
El
número de cromosomas se mantiene constante entre los individuos de
una misma especie.
Las
células somáticas de la mayoría de las plantas y animales son
diploides (tienen una dotación doble de cromosomas), mientras que
sus gametos son haploides (tienen una dotación simple). Las células
poliploides tienen más de dos dotaciones cromosómicas. El número
haploide de cromosomas se designa n y el número diploide, 2n.
En
toda célula diploide, cada cromosoma tiene su par homólogo. Uno de
los cromosomas homólogos proviene del gameto de uno de los
progenitores y su par, del gameto del otro progenitor.
La
meiosis: una reducción en el número de cromosomas
La
meiosis consiste en dos divisiones sucesivas que producen cuatro
células hijas haploides. De esta forma se compensa el efecto
multiplicador de la fecundación.

Durante
la meiosis, los miembros de cada par de cromosomas homólogos se
separan y cada gameto haploide (n), producido a partir de una célula
diploide (2n), lleva sólo un miembro de cada par. En la fecundación,
los núcleos del espermatozoide y del óvulo se unen en el cigoto,
cuyo núcleo contiene, nuevamente, los cromosomas homólogos de a
pares. Cada par está formado por un cromosoma homólogo proveniente
de un progenitor y el otro homólogo proveniente del otro progenitor.
En los diagramas usamos los colores rojo y verde para diferenciar los
cromosomas paternos de los maternos.
Al
comienzo de la meiosis I, los cromosomas homólogos se aparean y se
produce el entrecruzamiento: un fragmento de cromátida de un
homólogo se intercambia con un fragmento de cromátida del otro.
Durante la meiosis II, las cromátidas de cada homólogo se
distribuyen al azar entre las células hijas.
La
mitosis y la meiosis: procesos similares pero diferentes
En la meiosis, cada núcleo diploide se divide dos veces, pero los
cromosomas se duplican una sola. En la mitosis, en cambio, cada
división es precedida por una duplicación cromosómica.
Durante la meiosis, los cromosomas homólogos se aparean, se
entrecruzan y se segregan al azar. Nada de esto ocurre durante la
mitosis.
La mitosis ocurre en células haploides y diploides; la meiosis, sólo
en diploides.
La
meiosis en organismos con distintos ciclos vitales
En muchos organismos unicelulares y hongos haploides, la meiosis
ocurre inmediatamente después de la fusión de las células
fecundantes.
En las plantas que se reproducen en forma sexual se alternan una fase
haploide que por mitosis produce gametos y una fase diploide que por
meiosis produce esporas.
En los animales diploides, la meiosis produce gametos haploides.
Posibles
errores en la meiosis
Los cromosomas homólogos o sus cromátidas se pueden separar en
forma incorrecta y dar lugar a la aparición de gametos con
cromosomas faltantes o sobrantes.
Las
consecuencias de la reproducción sexual
En las especies con reproducción sexual ocurren tres procesos que
actúan como fuentes de variabilidad genética: el entrecruzamiento,
la segregación al azar de los cromosomas de los progenitores y la
fecundación. Esta variabilidad es un aspecto clave en el proceso
evolutivo de los seres vivos.
Fotosíntesis, luz y vida
Visión
general de la fotosíntesis: sus etapas
Los
organismos fotosintéticos productores de O2 usan energía lumínica,
CO2 y agua para producir la materia orgánica necesaria para su
alimentación. El O2 que liberan se forma con átomos provenientes
del agua.
La
fotosíntesis se realiza en dos etapas: la lumínica, en la que se
utiliza la energía de la luz para sintetizar ATP y NADPH, y la
fijadora de carbono, que utiliza los productos de la primera etapa
para la producción de azúcares.
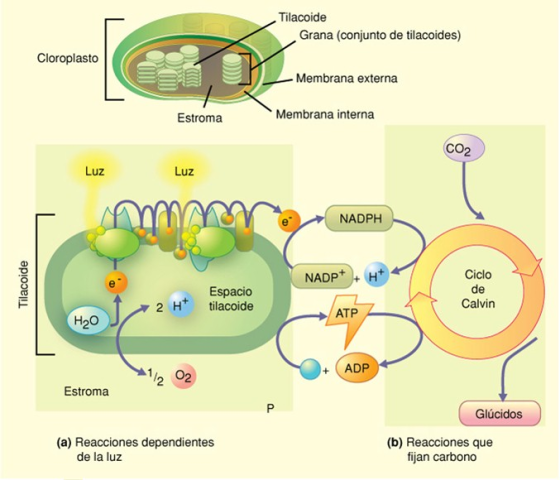
La
fotosíntesis ocurre en dos etapas: las reacciones dependientes de la
luz y las reacciones que fijan carbono. (a) En las reacciones
dependientes de la luz, la absorción de la energía lumínica por
las moléculas de clorofila a en la membrana del tilacoide inicia un
transporte de electrones y la formación de un gradiente de protones
a partir del cual se produce ATP. Durante este proceso, la molécula
de agua se escinde y se liberan moléculas de oxígeno gaseoso. Los
electrones son finalmente absorbidos por el NADP+ y se forma NADPH.
(b) En las reacciones que fijan carbono, que ocurren en la estroma
del cloroplasto, se sintetizan glúcidos a partir del CO2 y el
hidrógeno que transporta el NADPH. Este proceso utiliza la energía
del ATP y el NADPH producidos en la etapa dependiente de la luz y,
como veremos más adelante, implica una serie de reacciones que
constituyen el ciclo de Calvin.
La
fotosíntesis se realiza en los cloroplastos: los tilacoides
En
los eucariontes, la fotosíntesis se realiza en los cloroplastos,
organelas que poseen una membrana externa y otra interna. La membrana
interna rodea una solución densa, la estroma, donde se encuentran
las membranas tilacoides, que tienen forma de sacos aplanados
dispuestos en forma apilada. Las reacciones de la etapa lumínica
ocurren en los sacos tilacoides y las que fijan el carbono, en la
estroma.
Los
sacos tilacoides de los procariontes fotosintéticos pueden formar
parte de la membrana celular, estar aislados en el citoplasma o
constituir una estructura compleja de la membrana interna.
La
naturaleza de la luz
El
modelo ondulatorio de la luz permite a los físicos describir
matemáticamente ciertos aspectos de la luz y el modelo fotónico
permite otro tipo de cálculos y predicciones matemáticas. Estos dos
modelos ya no se consideran opuestos uno al otro, sino
complementarios, en el sentido de que es necesaria una síntesis de
ambos para una descripción completa del fenómeno que conocemos como
luz.
Los
sistemas vivos absorben la energía lumínica mediante el uso de
pigmentos. Los organismos fotosintéticos tienen distintos tipos de
pigmentos: la clorofila, que se encuentra en los sacos tilacoides,
los carotenoides y las ficobilinas. Existen diferentes tipos de
clorofila: la clorofila a, que colecta energía luminosa y está
involucrada en la transformación de energía lumínica en química;
la clorofila b, presente en las plantas y las algas verdes, y la
clorofila c de las algas marrones.
La
correspondencia entre el espectro de absorción de las clorofilas a y
b y el espectro de absorción de la fotosíntesis indica una estrecha
relación entre ésta y aquéllas (en ambos casos se observan dos
picos, uno en la zona del rojo y otro en la del azul). Los
carotenoides absorben en forma muy eficiente longitudes de onda que
no son absorbidas por la clorofila.
El
transporte de electrones: los fotosistemas y la ATP sintetasa
Los
organismos fotosintéticos poseen dos fotosistemas, cada uno formado
por una antena colectora de luz y un centro de reacción fotoquímico
que incluye una molécula de clorofila a. Ambos fotosistemas se
diferencian por el pico de absorción de la clorofila: el Fotosistema
I lo presenta a 700 nm; el Fotosistema II, a 680 nm.
En
un flujo no cíclico de electrones, los dos fotosistemas trabajan en
forma simultánea y continua. Así se produce un flujo permanente de
electrones desde el agua al Fotosistema II, de éste al Fotosistema I
y de este último al NADP+.
Durante
el transporte de electrones, los protones presentes en la estroma son
enviados al espacio intertilacoide, creando un gradiente cuya energía
se utiliza para sintetizar ATP. La síntesis de ATP a partir de
energía lumínica se conoce como fotofosforilación.
Cuando
los dos fotosistemas trabajan en forma independiente, se forma un
flujo cíclico de electrones. En este caso no se forma NADPH, pero se
sintetiza ATP. Es una ruta alternativa que permite regular la
cantidad de NADPH y ATP formados en presencia de luz y,
probablemente, aumenta la eficiencia en la formación de ATP cuando
coexiste con el flujo no cíclico de electrones.
Los fotosistemas:

La
energía lumínica atrapada en la molécula reactiva de la clorofila
a del Fotosistema II lanza los electrones a un nivel de energía
superior. Estos electrones son reemplazados en la molécula de
clorofila a por electrones que provienen indirectamente de moléculas
de agua que se escinden liberando además protones (H+) y gas
oxígeno. Los electrones pasan desde el aceptor de electrones
primario, a lo largo de una cadena de transporte de electrones, a un
nivel de energía inferior, el centro de reacción del Fotosistema I.
A medida que pasan a lo largo de esta cadena de transporte de
electrones, se forma un gradiente de protones a partir del cual se
sintetiza ATP. La energía lumínica absorbida por el Fotosistema I
lanza los electrones a otro aceptor primario. Desde este aceptor, los
electrones son transferidos mediante otros transportadores al NADP+ y
se forma NADPH. Los electrones eliminados del Fotosistema I son
reemplazados por los del Fotosistema II. El ATP y el NADPH
representan la ganancia neta de las reacciones que capturan energía.
La
fotofosforilación:
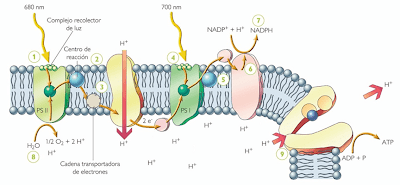
Moléculas
y complejos moleculares que participan de las reacciones directamente
dependientes de la luz. Entre ellos, se distinguen los pigmentos, los
transportadores de electrones, los Fotosistemas I y II y ciertas
enzimas como las ATP sintetasas. La disposición particular de estas
moléculas en la membrana tilacoide hace posible la síntesis
quimiosmótica del ATP durante la fotofosforilación. En este
proceso, los electrones de la molécula reactiva de clorofila a del
Fotosistema II son impulsados a niveles energéticos superiores por
la energía lumínica. A medida que descienden por una cadena de
transportadores de electrones hacia la molécula reactiva de
clorofila a del Fotosistema I, la energía que liberan se usa para
bombear protones (H+) desde la estroma al espacio tilacoide. Esto
crea un gradiente de protones. Cuando los protones se mueven a favor
del gradiente a través del complejo de la ATP sintetasa, desde el
espacio tilacoide a la estroma del cloroplasto, el ADP se fosforila a
ATP.
Las
reacciones que fijan carbono
El
ATP y el NADPH formados durante el transporte de electrones se
utilizan en la reducción del CO2 a glucosa. La incorporación de CO2
en compuestos orgánicos se conoce como fijación del carbono y
ocurre en forma cíclica (ciclo de Calvin). En las plantas verdes, el
CO2 llega a las células fotosintéticas a través de aberturas
especializadas llamadas estomas.
El
ciclo de Calvin comienza con la unión del CO2 a una molécula de
cinco carbonos (ribulosa bifosfato) que luego se divide en dos
moléculas de tres carbonos (fosfoglicerato). Cada seis vueltas del
ciclo se introducen seis moléculas de CO2 y se producen dos
moléculas de un azúcar de tres carbonos (gliceraldehído fosfato).
Las
plantas poseen un mecanismo de control que evita que el ciclo de
Calvin ocurra durante la noche. La luz lo estimula indirectamente y
las reacciones de fijación de carbono son inhibidas en la oscuridad.
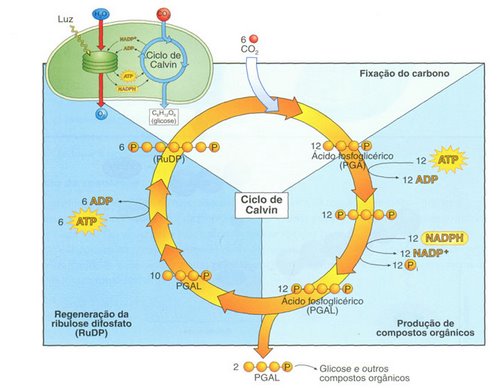
En
cada "vuelta" completa del ciclo ingresa una molécula de
CO2. Aquí se resumen seis ciclos, el número requerido para elaborar
dos moléculas de gliceraldehído fosfato, que equivalen a un azúcar
de seis carbonos. Se combinan seis moléculas de ribulosa bifosfato
(RuBP), un compuesto de cinco carbonos, con seis moléculas de CO2 y
se producen seis moléculas de un intermediario inestable que pronto
se escinde en doce moléculas de fosfoglicerato, un compuesto de tres
carbonos. Estos últimos se reducen a doce moléculas de
gliceraldehído fosfato. Diez de estas moléculas de tres carbonos se
combinan y se regeneran para formar seis moléculas de cinco carbonos
de RuBP. Las dos moléculas "extra" de gliceraldehído
fosfato representan la ganancia neta del ciclo de Calvin. Estas
moléculas son el punto de partida de numerosas reacciones que pueden
implicar, por ejemplo, la síntesis de glúcidos, aminoácidos y
ácidos grasos. La energía que impulsa al ciclo de Calvin proviene
del ATP y el NADPH producidos por las reacciones de captura de
energía en la primera etapa de la fotosíntesis.
La
fotorrespiración ocurre cuando la concentración de CO2 en la hoja
es baja en relación con la de O2. Consiste en la oxidación de la
ribulosa bifosfato, con formación de CO2 y agua. Es un proceso que
disminuye la eficiencia fotosintética de las plantas.
En
las células del mesófilo de las plantas C4, el CO2 se une a un
compuesto de tres carbonos (fosfoenolpiruvato), formando oxalacetato.
Este último se convierte en malato y pasa a zonas más profundas de
la hoja, donde libera CO2 que ingresa en el ciclo de Calvin. Este
proceso, que involucra gasto de energía, representa una adaptación
a las sequías y a intensidades lumínicas y temperaturas altas.

El
CO2 se fija primero en las células del mesófilo como ácido
oxalacético. La PEP carboxilasa, a diferencia de la RuBP
carboxilasa, es incapaz de incorporar O2. Aun con concentraciones muy
bajas de CO2 y en presencia de abundante O2, la enzima trabaja
rápidamente uniendo el CO2 al PEP. Comparada con la RuBP
carboxilasa, en presencia de O2 la PEP carboxilasa fija el CO2 más
rápido y en concentraciones menores, manteniendo baja la
concentración de CO2 dentro de las células cercanas a la superficie
de la hoja. El ácido oxalacético se transforma en ácido málico
que es transportado a las células de la vaina, donde libera CO2. El
CO2 así formado entra en el ciclo de Calvin. El ácido málico se
transforma en pirúvico que regresa a la célula del mesófilo, donde
es fosforilado a PEP.
En
plantas de ambientes secos existe una vía metabólica llamada
fotosíntesis CAM. La fijación de CO2 ocurre durante la noche y con
él se forma malato, que se almacena en las vacuolas. Durante el día,
el malato es liberado, se descarboxila y el CO2 ingresa en el ciclo
de Calvin.
Utilización
de los productos de la fotosíntesis
El
gliceraldehído fosfato producido por el ciclo de Calvin se integra
en glucosa o fructosa. Las células vegetales usan estas sustancias
para elaborar almidón, celulosa y sacarosa; las células animales
las usan para elaborar glucógeno. Todas las células utilizan
azúcares para la elaboración de otros carbohidratos, lípidos y
aminoácidos. Además, la oxidación del carbono fijado es la fuente
de energía del ATP en todas las células heterótrofas.
El
balance entre la fotosíntesis y la respiración
En
las plantas, la fotosíntesis y la respiración ocurren en forma
simultánea. La intensidad lumínica a la cual se igualan sus
velocidades es el punto de compensación para la luz. La
concentración de CO2 a la cual se igualan es el punto de
compensación para el CO2. Por debajo de estos puntos de
compensación, la respiración excede a la fotosíntesis y la planta
no crece. Como muchos órganos vegetales no fotosintetizan, para que
una planta se mantenga y crezca, la fotosíntesis debe exceder
largamente la tasa de respiración.
Suscribirse a:
Entradas
(
Atom
)